遗传
编辑遗传是亲代通过无性生殖或者有性生殖将性状特征传给后代,后代细胞或生物体获得亲代遗传信息的过程。通过遗传,个体之间的变异可以积累并通过自然选择导致物种进化。生物学中对遗传的研究就叫做遗传学。
1 概述编辑
在人类中,眼睛颜色是遗传特征的一个例子:个体可能从父母中的其中一个继承“棕色眼睛特征”。[1]遗传性状由基因控制,生物体基因组中的全套基因称为其基因型。[2]
生物体结构和行为的一整套可观察到的特征被称为其表现型。这些性状源于基因型与环境的相互作用。[3]因此,生物体表型的许多方面都不是遗传的。例如,晒黑的皮肤来自于人的表型和阳光之间的相互作用;[4]因此,晒黑的皮肤不会传给孩子。然而,由于基因型的差异,一些人比其他人更容易晒黑:[5]一个显著的例子是具有白化病遗传特征的人,他们根本不晒黑,对晒伤非常敏感。[6]
可遗传的特征被认为是通过DNA(一种编码遗传信息的分子)代代相传的。[2]DNA是一种包含四种类型的碱基的长聚合物,它们是可以互换的。一个特定DNA分子的碱基序列指定了遗传信息:这相当于一段文字的字母序列。[7]在细胞通过有丝分裂进行分裂之前,DNA被复制,因此产生的两个细胞中的每一个都将继承DNA序列。DNA分子中指定单个功能单元的部分称为基因;不同的基因有不同的碱基序列。在细胞内,DNA的长链形成称为染色体的浓缩结构。生物体以同源染色体的形式从亲代那里遗传到遗传物质,其中含有编码基因的DNA序列的独特组合。染色体中DNA序列的特定位置称为基因座。如果某一特定位点上的DNA序列在不同个体之间存在差异,则该序列的不同形式称为等位基因。DNA序列可以通过突变而改变,产生新的等位基因。如果基因发生突变,新的等位基因可能会影响基因控制的性状,从而改变生物体的表型。[8]
然而,虽然等位基因和性状之间的这种简单对应在某些情况下有效,但大多数性状更复杂,并由生物体内部和生物体之间的多个相互作用基因控制。[9][10]发育生物学家认为,遗传网络和细胞间通讯的复杂相互作用可能导致遗传变异,这可能是发育可塑性和导管化机制的基础。[11]
最近的发现证实了一些重要的遗传变化的例子,这些变化不能用DNA分子的直接作用来解释。这些现象被归类为表观遗传系统,是由基因引起或独立进化而来的。对表观遗传的模式和机制的研究仍处于科学的起步阶段,然而,这一领域的研究吸引了最近的许多活动,因为它拓宽了遗传力和进化生物学的范围。[12] DNA甲基化标记染色质,自我维持的代谢循环、 RNA干扰导致的基因沉默,蛋白质的三维构象(如朊病毒)是在生物体水平上发现表观遗传系统的领域。[13][14]遗传性也可能在更大的范围内发生。例如,生态位构建过程中的生态遗传被定义为生物体在其环境中的规律性和重复性活动。这就产生了一种影响,这种影响会修改并反馈给后代的选择机制。后代遗传基因和祖先生态行为所产生的环境特征。[15]在进化中不受基因直接控制的遗传力的其他例子包括文化性状、群体遗传力和内共生的遗传。[16][17][18]这些在基因之上起作用的遗传性的例子被广泛地覆盖在多级或分级选择的标题之下,这在进化科学史上一直是一个辩论激烈的主题。[17][19]
2 与进化论的关系编辑
当查尔斯·达尔文在1859年提出他的进化理论时,它的主要问题之一是缺乏遗传的潜在机制。[20]达尔文相信混合遗传和后天性状遗传的结合(泛生)。混合遗传只会在几代内导致种群间的一致性,然后消除自然选择作用的种群中的变异。[21]这导致达尔文在《物种起源》的后期版本和他后期的生物学著作中采用了一些拉马克思想。[22]达尔文研究遗传的主要方法是概述它是如何工作的(注意到在繁殖时亲代没有明确表达的特征可以遗传,某些特征可能是与性别有关的,等等),而不是提出机制。
达尔文最初的遗传模型被他的堂兄弟弗朗西斯·高尔顿采用,然后被他大幅度修改,他为生物统计遗传学派奠定了框架。[23]高尔顿没有找到证据来支持达尔文的泛生模型,该模型依赖于后天获得的特性。[24]
19世纪80年代,奥古斯特·魏斯曼(August Weismann)剪掉了几代小鼠的尾巴,发现它们的后代继续发育出尾巴,这表明后天性状几乎不遗传。[25]
3 历史编辑
古代的科学家对遗传有各种各样的想法:提奥夫拉斯图斯提出雄花导致雌花成熟;[26] 希波克拉底推测,“种子”是由身体的各个部位产生的,并在受孕时传给后代;[27]亚里士多德认为男性和女性的体液在受孕时是混合的。[28] 公元前458年,埃斯库罗斯提议由男性来养育后代,而女性则是“她所播种的年轻生命的保姆”。[29]
古代对遗传的理解在18世纪转变为两种有争议的学说。表观生成学说和预形成学说是理解遗传的两种截然不同的观点。亚里士多德的表观发生学说声称胚胎是不断发育的。父母特征的改变会在胚胎存续期间被传递。这一学说的基础是基于后天性状遗传理论。与此直接对立的是,预形成学说声称“相似产生相似”,即细菌会进化产生与父母相似的后代。预格式化主义者认为生育是揭示很久以前就已经被创造出来的东西的行为。然而,这一点受到了19世纪细胞理论的质疑,在该理论中,生命的基本单位是细胞,而不是生物体的某些预先形成的部分。还设想了各种遗传机制,包括融合遗传,但没有进行适当的测试或量化,所以后来有争议。尽管如此,人们还是能够通过人工选择发展家畜品种和农作物。后天性状的遗传也是早期拉马克进化论的一部分。
18世纪,荷兰显微镜学家安东尼·范·列文虎克(1632-1723)在人类和其他动物的精子中发现了“微生物”。[30]一些科学家推测他们在每个精子中看到了一个“小人”(人造人)。这些科学家形成了一个被称为“精子学家”的思想流派。他们认为女性对下一代的唯一贡献是人造人成长的子宫和子宫的产前影响。[31]另一个相反的学派,产卵学家,认为未来的人类在卵子中,精子仅仅刺激卵子的生长。产卵学家认为妇女携带含有男孩和女孩的卵,并且后代的性别早在受孕前就已经确定了。[32]
3.1 孟德尔:遗传学之父
基因微粒遗传的想法可以归因于摩拉维亚 [33]蒙克·孟德尔,他于1865年发表了关于豌豆植物的著作。然而,他的作品没有多少人知道,并于1901年被重新发现。起初人们认为,孟德尔定律只解释了很大的(定量)差异,例如孟德尔在他的豌豆植株中看到的那些差异 –直到 r . a . Fisher (1918)的论文“孟德尔定律假设下的亲属之间的相关性”孟德尔的总体贡献给了科学家们一个有用的概述,即性状是可遗传的,基因(数量)的加法效应的想法才被实现。他的豌豆植物演示成为孟德尔性状研究的基础。这些特征可以在一个单一的位点上找到。[34]
3.2 遗传学和遗传的现代发展
20世纪30年代,费希尔和其他人的工作将孟德尔学派和生物特征学派结合起来,形成了现代综合理论。现代合成技术填补了实验遗传学家和自然学家之间的空白;以及古生物学家之间的联系:[35][36]
- 所有的进化现象都可以用一种与已知的遗传机制和自然学家的观察证据相一致的方式来解释。
- 进化是渐进的:小的遗传变化,自然选择的重组。物种(或其他分类单元)之间的不连续被解释为通过地理分离和灭绝(而不是跃移)逐渐产生的。
- 绝大多数情况下,选择是改变的主要机制;即使是微小的优势也很重要。选择的对象是其周围环境的表型。遗传漂变的作用是模棱两可的;虽然最初得到了多勃赞斯基的大力支持,但后来随着生态遗传学的结果的获得而被降级。
- 群体思维的首要性:自然群体的遗传多样性是进化的关键因素。自然选择在野外的力量比预期的要大;生态位占有等生态因素的影响和基因流动障碍的意义都很重要。
关于物种形成发生在种群繁殖隔离之后的观点一直备受争议。[37]在植物中,多倍体必须包含在任何物种形成的观点中。诸如“进化主要由一代人和另一代人之间等位基因的频率的变化组成”等表述是在相当晚的时候提出的。传统观点认为发育生物学(“evo-devo ”)在合成过程中几乎没有发挥作用,但是史蒂芬·杰·古尔德对加文·德·比尔工作的描述表明他可能是一个例外。[38]
几乎合成上的各方面都受到挑战,却又都取得了不同程度的成功。然而,毫无疑问,这种合成是进化生物学的一个伟大里程碑。[39]它澄清了许多困惑,并直接促进了二战后时代的大量研究。
然而,当特罗菲姆·李森科强调了拉马克主义者关于后天性状遗传的思想时,在苏联引起了现在被称为李森科主义的强烈反对。这场运动影响了农业研究,导致了20世纪60年代的粮食短缺,并严重影响了苏联。[40]
越来越多的证据表明,人类[41]和其他动物的表观遗传存在跨代遗传。[42]
3.3 常见遗传疾病
-
- 唐氏综合征
- 镰状细胞病
- 苯丙酮尿症 (PKU)
- 血友病[34]
4 类型编辑
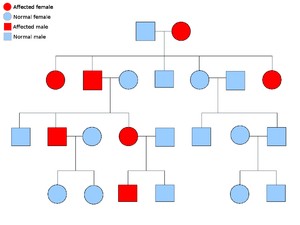
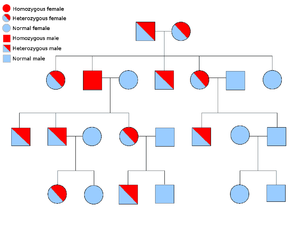
4.1 显性和隐性等位基因
如果一个等位基因总是在生物体的外观(表型)中表达,那么它就被认为是显性的,前提是它的至少一个拷贝是存在的。例如,在豌豆中,绿色豆荚的等位基因G,比黄色豆荚的等位基因g占主导地位。因此,具有这对等位基因的豌豆植株,无论是GG(纯合子)还是Gg(杂合子)都会有绿色的豆荚。黄色豆荚的等位基因是隐性的。只有当等位基因存在于两条染色体中时,这种等位基因的影响才会显现出来,即gg(纯合体)。
生物遗传模式的描述包括三个主要类别:
-
1.涉及的基因座数量
- 单基因(也称为“简单”)—一个基因座
- 寡遗传–基因座很少
- 多基因–基因座很多
- 2.涉及染色体
-
- 常染色体–基因座不位于性染色体上
- 性染色体-基因座位于性染色体上
- X染色体基因座位于 X染色体上(更常见的情况)
- Y染色体基因座位于 Y染色体上
- 线粒体–基因座位于线粒体DNA 上
-
3.相关性基因型–表现型
- 显性
- 中间体(也称为“密码子”)
- 隐性
- 超显性
- 显性不足
这三个类别是按照上述顺序对继承模式的每个精确描述的一部分。此外,还可以按以下方式添加更多规范:
- 4.偶然因素和环境的相互作用
-
- 外显率
- 完全外显
- 不完全外显(百分比数)
- 表现度
- 不变的
- 可变的
- 遗传力(在多基因遗传中,有时也在寡基因遗传模式中)
- 母系或父系的印迹现象(也见表观遗传学)
- 外显率
- 5.与性别有关的相互作用
-
- 性别相关遗传(性染色体基因座)
- 性别受限的表型表达(例如,隐睾)
- 通过母系遗传(在线粒体DNA 基因座的情况下)
- 通过父系遗传(在 Y染色体位点的情况下)
-
6.位点-位点相互作用
- 异位显性与其他基因座(例如,超显性)
- 基因耦合与其他基因座(另请参见穿越)
- 纯合致死因子
- 半致死因子
遗传模式的确定和描述也主要是通过系谱数据的统计分析来实现。在所涉及的基因座是已知的情况下,也可以采用分子遗传学的方法。
参考文献
- [1]
^Sturm RA; Frudakis TN (2004). "Eye colour: portals into pigmentation genes and ancestry". Trends Genet. 20 (8): 327–332. doi:10.1016/j.tig.2004.06.010. PMID 15262401..
- [2]
^Pearson H (2006). "Genetics: what is a gene?". Nature. 441 (7092): 398–401. Bibcode:2006Natur.441..398P. doi:10.1038/441398a. PMID 16724031..
- [3]
^Visscher PM; Hill WG; Wray NR (2008). "Heritability in the genomics era – concepts and misconceptions". Nat. Rev. Genet. 9 (4): 255–266. doi:10.1038/nrg2322. PMID 18319743..
- [4]
^Shoag J; et al. (Jan 2013). "PGC-1 coactivators regulate MITF and the tanning response". Mol Cell. 49 (1): 145–157. doi:10.1016/j.molcel.2012.10.027. PMC 3753666. PMID 23201126..
- [5]
^Pho LN; Leachman SA (Feb 2010). "Genetics of pigmentation and melanoma predisposition". G Ital Dermatol Venereol. 145 (1): 37–45. PMID 20197744..
- [6]
^Oetting WS; Brilliant MH; King RA (1996). "The clinical spectrum of albinism in humans and by action". Molecular Medicine Today. 2 (8): 330–335. doi:10.1016/1357-4310(96)81798-9. PMID 8796918..
- [7]
^Griffiths, Anthony, J.F.; Wessler, Susan R.; Carroll, Sean B.; Doebley J (2012). Introduction to Genetic Analysis (10th ed.). New York: W.H. Freeman and Company. p. 3. ISBN 978-1-4292-2943-2..
- [8]
^Futuyma, Douglas J. (2005). Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc. ISBN 978-0-87893-187-3..
- [9]
^Phillips PC (2008). "Epistasis – the essential role of gene interactions in the structure and evolution of genetic systems". Nat. Rev. Genet. 9 (11): 855–867. doi:10.1038/nrg2452. PMC 2689140. PMID 18852697..
- [10]
^Wu R; Lin M (2006). "Functional mapping – how to map and study the genetic architecture of dynamic complex traits". Nat. Rev. Genet. 7 (3): 229–237. doi:10.1038/nrg1804. PMID 16485021..
- [11]
^Jablonka, E.; Lamb, M.J. (2002). "The changing concept of epigenetics" (PDF). Annals of the New York Academy of Sciences. 981 (1): 82–96. Bibcode:2002NYASA.981...82J. doi:10.1111/j.1749-6632.2002.tb04913.x. PMID 12547675. Archived from the original (PDF) on 2011-05-11..
- [12]
^Jablonka, E.; Raz, G. (2009). "Transgenerational epigenetic inheritance: Prevalence, mechanisms, and implications for the study of heredity and evolution" (PDF). The Quarterly Review of Biology. 84 (2): 131–176. CiteSeerX 10.1.1.617.6333. doi:10.1086/598822. PMID 19606595..
- [13]
^Bossdorf, O.; Arcuri, D.; Richards, C.L.; Pigliucci, M. (2010). "Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana". Evolutionary Ecology. 24 (3): 541–553. doi:10.1007/s10682-010-9372-7..
- [14]
^Jablonka, E.; Lamb, M. (2005). Evolution in four dimensions: Genetic, epigenetic, behavioural, and symbolic. MIT Press. ISBN 978-0-262-10107-3..
- [15]
^Laland, K.N.; Sterelny, K. (2006). "Perspective: Seven reasons (not) to neglect niche construction" (PDF). Evolution. 60 (8): 1751–1762. doi:10.1111/j.0014-3820.2006.tb00520.x. Archived from the original (PDF) on 2011-08-19..
- [16]
^Chapman, M.J.; Margulis, L. (1998). "Morphogenesis by symbiogenesis" (PDF). International Microbiology. 1 (4): 319–326. PMID 10943381. Archived from the original (PDF) on 2014-08-23..
- [17]
^Wilson, D. S.; Wilson, E.O. (2007). "Rethinking the theoretical foundation of sociobiology" (PDF). The Quarterly Review of Biology. 82 (4): 327–348. doi:10.1086/522809. PMID 18217526. Archived from the original (PDF) on 2011-05-11..
- [18]
^Bijma, P.; Wade, M.J. (2008). "The joint effects of kin, multilevel selection and indirect genetic effects on response to genetic selection". Journal of Evolutionary Biology. 21 (5): 1175–1188. doi:10.1111/j.1420-9101.2008.01550.x. PMID 18547354..
- [19]
^Vrba, E.S.; Gould, S.J. (1986). "The hierarchical expansion of sorting and selection: Sorting and selection cannot be equated" (PDF). Paleobiology. 12 (2): 217–228. doi:10.1017/S0094837300013671..
- [20]
^Griffiths, Anthony, J.F.; Wessler, Susan R.; Carroll, Sean B.; Doebley, John (2012). Introduction to Genetic Analysis (10th ed.). New York: W.H. Freeman and Company. p. 14. ISBN 978-1-4292-2943-2..
- [21]
^Charlesworth, Brian & Charlesworth, Deborah (November 2009). "Darwin and Genetics". Genetics November 2009 No. 3 757–766. 183 (3): 757–766. doi:10.1534/genetics.109.109991. PMC 2778973. PMID 19933231..
- [22]
^Bard, Jonathan BL (2011). "The next evolutionary synthesis: from Lamarck and Darwin to genomic variation and systems biology". Cell Communication and Signaling. 9 (30): 30. doi:10.1186/1478-811X-9-30. PMC 3215633. PMID 22053760..
- [23]
^"Francis Galton (1822-1911)". Science Museum. Retrieved March 26, 2013..
- [24]
^Liu Y. (May 2008). "A new perspective on Darwin's Pangenesis". Biol Rev Camb Philos Soc. 83 (2): 141–149. doi:10.1111/j.1469-185X.2008.00036.x. PMID 18429766..
- [25]
^Lipton, Bruce H. (2008). The Biology of Belief: Unleashing the Power of Consciousness, Matter and Miracles. Hay House, Inc. p. 12. ISBN 978-1-4019-2344-0..
- [26]
^Negbi, Moshe (Summer 1995). "Male and female in Theophrastus's botanical works". Journal of the History of Biology. 28 (2): 317–332. doi:10.1007/BF01059192..
- [27]
^Hipócrates (1981). Hippocratic Treatises: On Generation – Nature of the Child – Diseases Ic. Walter de Gruyter. p. 6. ISBN 978-3-11-007903-6..
- [28]
^"Aristotle's Biology – 5.2. From Inquiry to Understanding; from hoti to dioti". Stanford University. Feb 15, 2006. Retrieved March 26, 2013..
- [29]
^欧墨尼得斯658-661.
- [30]
^Snow, Kurt. "Antoni van Leeuwenhoek's Amazing Little "Animalcules"". Leben. Archived from the original on April 24, 2013. Retrieved March 26, 2013..
- [31]
^Lawrence, Cera R. (2008). Hartsoeker's Homunculus Sketch from Essai de Dioptrique. Embryo Project Encyclopedia. ISSN 1940-5030. Archived from the original on 2013-04-09. Retrieved March 26, 2013..
- [32]
^Gottlieb, Gilbert (2001). Individual Development and Evolution: The Genesis of Novel Behavior. Psychology Press. p. 4. ISBN 978-1-4106-0442-2..
- [33]
^Henig, Robin Marantz (2001). The Monk in the Garden : The Lost and Found Genius of Gregor Mendel, the Father of Genetics. Houghton Mifflin. ISBN 978-0-395-97765-1. The article, written by an obscure Moravian monk named Gregor Mendel.
- [34]
^尼尔·卡尔森(2010年)。心理学:行为科学,第206页。多伦多:加拿大皮尔逊。ISBN 978-0-205-64524-4。OCLC 1019975419.
- [35]
^迈尔&普罗文1998.
- [36]
^1982年5月。生物思维的成长:多样性、进化与继承。哈佛,坎波斯。第567页及其后各页。.
- [37]
^Palumbi, Stephen R. (1994). "Genetic Divergence, Reproductive Isolation, and Marine Speciation". Annual Review of Ecology and Systematics. 25: 547–572. doi:10.1146/annurev.es.25.110194.002555..
- [38]
^古尔德S.J .个体发育和系统发育。哈佛大学,1977年。第221-222页.
- [39]
^Handschuh, Stephan; Mitteroecker, Philipp (June 2012). "Evolution – The Extended Synthesis. A research proposal persuasive enough for the majority of evolutionary biologists?". The International Society for Human Ethology. 27 (1–2): 18–21. ISSN 2224-4476..
- [40]
^Harper, Peter S. (2017-08-03). "Human genetics in troubled times and places". Hereditas. 155: 7. doi:10.1186/s41065-017-0042-4. ISSN 1601-5223. PMC 5541658. PMID 28794693..
- [41]
^Szyf, M (2015). "Nongenetic inheritance and transgenerational epigenetics". Trends in Molecular Medicine. 21 (2): 134–144. doi:10.1016/j.molmed.2014.12.004. PMID 25601643..
- [42]
^Kishimoto, S; et al. (2017). "Environmental stresses induce transgenerationally inheritable survival advantages via germline-to-soma communication in Caenorhabditis elegans". Nature Communications. 8: 14031. Bibcode:2017NatCo...814031K. doi:10.1038/ncomms14031. hdl:2433/217772. PMID 28067237..
暂无